Theories
|

|
"Summarized in a few words, the prime cause of cancer is the replacement of the respiration of oxygen in
the normal body cells by a fermentation of sugar."
- Otto Heinrich Warburg.
The prevailing theory on cancer is that it is a genetic disease, which goes something like this:
With rare exceptions, each human cell contains a nucleus housing some 25,000 genes; some are oncogenes (genes with the potential to take the cell on the road to cancer) and some are tumour suppressor genes (genes that protect the cell from one step on the road to cancer). Spontaneous, permanent alterations in genes, or mutations, occur at a slow rate all the time, but when enough mutations occur in enough oncogenes and tumour suppressor genes, the ability of the cell to genetically regulate its growth is destroyed and an abnormal mass of tissue, or tumour, develops. Eventually, a subset of tumour cells acquires a set of mutations that grants the ability to metastasize (detach themselves from the primary tumour, enter the circulation, evade immune attack, exit the circulation at a distant site, and invade and proliferate in a distant organ).
Although intriguing, this genetic theory of cancer is not consistent with several key facts:
(1) Cybrid experiments - A cybrid is a cell that contains the nucleus of one cell and the mitochondria of another cell (the nucleus is an organelle that contains most of the cell's genes, the mitochondria are organelles that produce most of the cell's energy). Repeated experiments from multiple labs have shown that a cancer cell nucleus combined with normal mitochondria produces a normal cell, but cancer cell mitochondria combined with a normal cell nucleus produces a cancer cell (1-3). These rather striking results strongly imply that a cell's predisposition towards cancer is dependent on its energy-producing mitochondria, not its gene-containing nucleus.
|
In any animal cell, the nucleus contains most of the cell's genes whereas the more numerous mitochondria produce most of the cell's energy. Cybrid experiments imply that cancer is initiated by the mitochondria. |
(2) Slow spontaneous mutation rate - According to the genetic theory of cancer, spontaneous mutations are the ultimate source of genetic damage in a cell. Yet by the time a tumour has formed, it has already developed hundreds or even thousands of mutations, and the mutations differ from one cancer cell to the next within most tumours (4-6). The spontaneous mutation rate is far too slow to produce so many different mutations over the course of a few years, or even a lifetime. Some advocates of the genetic theory try to counter this fact by suggesting that many of the mutations first occur in caretaker genes (genes that stabilize other genes, protecting them against mutations) such as p53, which results in a faster mutation rate in the remaining genes, but this argument begs the question as to why nature would make caretaker genes so prone to mutation in the first place.
(3) Metastasis - Metastasis is an extremely complex cell behaviour that allows cells to detach themselves from the primary tumour, travel to a distant tissue or organ while evading the host's multi-layered defence system, and settle in that tissue or organ. According to the genetic theory of cancer, such behaviour requires that the cancerous cell obtain a diverse array of beneficial survival tricks from random mutations - and yet, it is a fundamental tenet of evolution that random mutations are virtually always deleterious to cells (7). Thus, the genetic theory presumes that cancer cells "get lucky" on a suspiciously high number of occasions, acquiring numerous beneficial mutations at random that enable them to undergo such complex behaviour - consistently, from one cancer patient to the next.
|
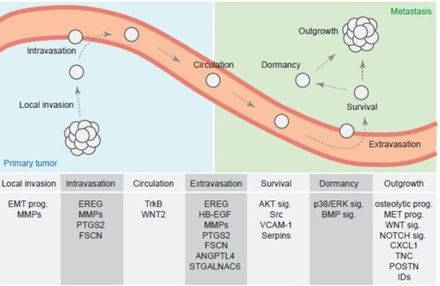
The steps involved in metastasis (plus genes and signalling pathways). It seems a tad far-fetched to believe that cancer cells get lucky in randomly acquiring all these benefical mutations that endow them with an extremely complex behaviour - consistently, in millions of cancer patients. |
In addition to these unsettling truths, the genetic theory of cancer only attempts to explain the "how" of cancer, without addressing the "why" of cancer. This is crucial; consider the example of an alcoholic. Although we know that a man's alcoholism results from excessive alcohol consumption, and we may talk at length about how alcohol affects the human body, this mechanistic explanation does not tell us why the man continues to drink alcohol. Did he suffer a recent tragedy? Does he have an addictive personality? What about his environment, does he live right next to a liquor store? These factors may all be relevant to his condition. Without understanding the why of this man's alcoholism, we'll never cure it.
Returning to cancer, in analogous fashion we can talk about mutated genes and other cell defects until we turn blue in the face, but this does not tell us why cancer evolved in the first place. Does cancer play a role in evolution? Are there circumstances where it is beneficial for a cell to become cancerous? Does the cancer cell environment matter? Just like the alcoholic, without understanding why cancer exists, we'll never cure it.
A longstanding criticism of the field of cancer biology is that it does not comprehensively explain why cancer arose in evolution (8). Some cancer researchers advocate the rogue cell hypothesis, which posits that cancer cells are essentially selfish, evolving their array of survival tricks in the face of selective pressures from their host's defenses (7,9). Yet this makes little evolutionary sense, for as soon as the cancer cells win over their host, their host dies - and the cancer cells with it.
This is the state of the cancer field today. The genetic theory attempts to explain how cancer is initiated, but its explanation is not consistent with several critical facts. Moreover, cancer biology lacks a comprehensive evolutionary theory explaining why cancer exists at all.
If we want to cure cancer, we need a how that fits the facts, and we need a why that makes evolutionary sense.
First, the how.
Cancer - The How
Let's drift away from genetics, take a look at some facts about how cells function - specifically, how they metabolize.
Metabolism describes all the chemical reactions within a cell, with the six-carbon molecule glucose (the building block of carbohydrates) as the star of the show. For the cell to produce energy, glucose must enter a series of reactions in the cell's cytoplasm called glycolysis, producing a molecule called pyruvate. Depending on oxygen levels outside the cell, pyruvate can go one of two ways:
(1) Fermentation - If oxygen levels are low, pyruvate is converted to lactate in the cells' cytoplasm, resulting in relatively modest amounts of energy produced.
(2) Respiration - If oxygen levels are high, pyruvate is transported into the cell's mitochondria where it enters a series of reactions called the Krebs cycle and electron transport chain, with a lot more energy produced.
Normal human cells are metabolically flexible depending on oxygen levels outside the cell; they can adjust their metabolism to ferment or respire as needed. If oxygen levels are high, normal cells undergo the Pasteur Effect (suppress fermentation in favour of respiration); this is advantageous, since respiration produces much more energy per glucose molecule compared to fermentation. It is only when oxygen levels become too low for efficient respiration that normal cells resort to fermentation as their primary means of energy production.
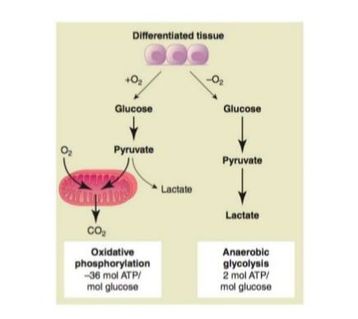
|
|
In contrast, cancer cells are metabolically inflexible regardless of oxygen levels outside the cell. Even in the presence of high oxygen levels, cancer cells undergo the Warburg Effect (ferment in the presence of oxygen), in which they crank up their rates of glycolysis to five to ten (or even over a hundred) times the normal rate, with much of the extra pyruvate converted into lactate (10). No matter how much oxygen is available, cancer cells ferment more, respire less. Why do cancer cells insist on doing this, when respiration produces much more energy per glucose molecule? It seems counter-productive. However, it may be that they simply don't have a choice - it has been argued that the bulk of experimental evidence indicates that virtually all cancer cells have dysfunctional mitochondria, thus their ability to respire is impaired, so they have no other option but to ferment (11).
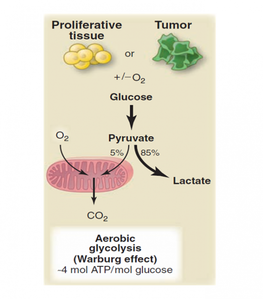
|
|
Normal cells also adjust their metabolism in response to nutrient levels outside the cell; for example, they respond to glucose via the Crabtree Effect (high glucose levels inhibit respiration), but they also respond to other organic molecules such as amino acids (the building blocks of proteins) and lipids (building blocks of fats). In a more general sense, when nutrient levels are high, the cell adjusts its metabolism towards anabolism (using ATP energy to build up the cell; promotes cell growth and replication), and when nutrients are limited, the cell adjusts its metabolism towards catabolism (recycling cell parts to create ATP energy; promotes cell survival). How does the cell gauge the nutrient levels in its environment? It uses hormones and enzymes that act as nutrient sensors (12), the three main ones being:
(1) Insulin - Insulin is an anabolic hormone and its levels are strongly increased by glucose; it is also moderately increased by amino acids and even slightly increased by lipids. Insulin is the key regulator of insulin-like growth factor (IGF-1), which promotes further anabolism.
(2) mTOR - An enzyme that drives cell survival, growth, and proliferation. Like insulin, mTOR is also anabolic and is strongly increased by amino acids. High mTOR activity inhibits autophagy, a catabolic process that recycles worn-out cell components so they can be used for energy. If a normal cell has acquired enough damage, autophagy can result in "cell suicide" which can aid the survival of the entire organism.
(3) AMPK - An enzyme that acts as a reverse sensor of all nutrients. AMPK is catabolic and therefore acts in the opposite direction of insulin and mTOR, decreasing in response to glucose, amino acids, and lipids. High AMPK levels directly and indirectly stimulate autophagy, thus promoting the recycling of worn-out cell components for energy.
In contrast to normal cells, cancer cells are less able to flip back and forth between anabolism and catabolism. Regardless of nutrient availability, they are "addicted" to nutrients, especially glucose and the amino acid glutamine (11,13). Although they ferment glucose via the Warburg Effect to create ATP energy, many cancer cells also metabolize glutamine via the Krebs cycle in a sort of "pseudo-respiration" that may provide much of their energy requirements; some tumours are even more dependent on glutamine for their energy needs than they are on glucose (11,13,14). Using glucose and glutamine to fuel their metabolism, cancer cells ignore many extracellular signals that instruct them to behave like normal cells; instead, they make critical cellular decision in a cell-autonomous manner (15), heedlessly upregulating their insulin receptors, upregulating mTOR, and downregulating AMPK (12). Moreover, while much of the pyruvate formed from the cancer cell's souped-up glycolysis is used to ferment ATP energy, much of the pyruvate is also diverted towards building an abundance of organic molecules (such as amino acids and lipids) that facilitate the enhanced, unrestricted growth and replication of the cell (15). Thus, cancer cells, with their insatiable thirst for glucose and glutamine, are "reprogrammed" into a permanent state of anabolism (15,16) - a state in which they constantly upregulate their insulin receptors, upregulate mTOR, and downregulate AMPK.
Given these facts, an alternative explanation that places less emphasis on cancer genetics and more emphasis on cancer metabolism can be formulated. It goes something like this:
Cancer originates with mitochondrial dysfunction, which can be due to sources such as toxins and radiation (plus others), with longer exposure times resulting in more dysfunction. This mitochondrial dysfunction impairs cell respiration but leaves fermentation intact; to maintain energy production the cell is forced to increase glycolysis and permanently undergo the Warburg Effect - ferment more, respire less. The pseudo-respiration of the amino acid glutamine in the Krebs cycle also significantly contributes to energy production. The Warburg Effect results in an excess of pyruvate, and that which is not fermented is directed towards growth and replication such that the cell remains in a permanent state of anabolism, upregulating its insulin receptors, upregulating mTOR, and downregulating AMPK. The cell is now cancerous - it cannot stop itself from constantly fermenting, growing, and replicating, resulting in the development of a tumour.
Note that this "mitochondrial dysfunction" theory of cancer has not explained metastasis yet, but bear with me.
Now, the why.
Cancer - The Why
Let's take a step back, see the bigger picture.
Evolution describes a process of natural selection by which traits that enhance survival and reproduction become more common over successive generations. Given its massive influence in biology, a plausible theory of cancer should make sense in an evolutionary context.
Life is 3-4 billion years old (17); it first appeared under an early atmosphere containing very little free oxygen, in primitive oceans awash with organic molecules. Early life started out as unicellular, but at some point it transitioned to close-knit colonies of partially differentiated cells displaying a rudimentary degree of cooperation and self-organization (7). Given that the atmosphere was lacking in oxygen, simple multicellular lifeforms had no choice but to constantly ferment, and given that the ocean was full of organic nutrients, the optimal survival and reproduction strategy would have been to remain anabolic so as to grow and reproduce as quickly as possible. Kind of like cancer cells.
|

Primitive oceans. Note the lack of atmospheric oxygen and abundance of oceanic nutrients. The optimal evolutionary strategy for cells in such an environment would be to constantly ferment, grow, and replicate. |
Roughly 1 billion years later, the appearance of cyanobacteria resulted in rising atmospheric oxygen levels (18). As free oxygen become readily available, simple multicellular lifeforms slowly gained the ability to respire, resulting in a greater capacity for energy production and the emergence of more complex multicellular lifeforms. Given the high oxygen availability and rising competition for nutrients, the optimal survival and reproduction strategy for complex multicellular lifeforms would have been to respire most of their energy needs while retaining a capacity to flip between anabolism and catabolism depending on whether environmental conditions favoured reproduction or survival. Like normal human cells.
In this scenario, the "newer" genes (such as tumour suppressor genes) that evolved with complex multicellular lifeforms were overlain on the "older" genes (such as oncogenes) that controlled simple multicellular lifeforms (7). However, the genetic code for simple multicellular lifeforms did not disappear, it was just suppressed - dormant, but intact. Indeed, the newer genes that evolved with complex multicellularity are the same ones that malfunction in cancer, implying that when things go wrong in the cells of complex multicellular lifeforms, some cells fall back to the strategy that successfully allowed their ancestors to survive and thrive over hundreds of millions of years - an ancient evolutionary strategy of non-stop fermentation, growth, and replication.
Seen this way, cancer is not a new condition, but a very old one that occurs when cell damage unlocks an ancient "toolkit" of pre-existing adaptations, re-establishing the dominance of an earlier layer of genes that controlled loose-knit colonies of partially differentiated cells (7). This evolutionary theory of cancer suggests that when things go wrong with the cells of a complex multicellular lifeform, the ancient genes in some of these cells reassert themselves, resulting in a subset of cells that behave like simple multicellular lifeforms...cells that we call cancer.
Let's examine the validity of this "ancient gene reactivation" theory of cancer:
(1) If true, cancer should exist far back in evolution - Indeed, cancer exists in all complex multicellular organisms examined, with the possible exception of the naked mole rat (7,19). The fossil record is strewn with examples of tumours occurring far back into evolution - for example, dinosaur tumours have been documented many times (20).
(2) If true, oncogenes should exist far back in evolution - Indeed, oncogenes are ancient in evolutionary terms - for example, the myc oncogene is probably the most frequently amplified oncogene in all of cancer (13), and it is at least 600 million years old (21), with newer studies revealing even older dates (22).
(3) If true, there should be other examples of failure of genetic suppression - Indeed, the genetic code of a multicellular lifeform contains the information for most or all of its evolutionary ancestors - for example, humans are sometimes born with excessive body hair reminiscent of our primate ancestors, as well as webbed feet and tails reminiscent of our even more distant ancestors (23). If the ancient genes for these traits exist somewhere in the human genome, so can others.
Moreover, this ancient gene reactivation theory of cancer provides a plausible explanation for the complex behaviour of cancer cells when they metastasize. When viewed as an unlocking of an ancient toolkit of pre-existing adaptations, the many beneficial mutations that occur in metastatic cancer cells no longer seem so "lucky." Rather, metastasis results from the reactivation of a genetic program of cell survival tricks, a program that evolved over hundreds of millions of years in simple multicellular lifeforms and was always present, but was later suppressed during the evolution of complex multicellular lifeforms. But was always present.
Let's put this all together.
A Metabolic Theory Of Cancer
Combining the two theories above results in a metabolic theory of cancer in which mitochondrial dysfunction results in an energy crisis forcing the cell to ferment, grow, and replicate constantly, followed by the reactivation of ancient genes that further instruct the cell to ferment, grow, and replicate constantly. These two theories are complementary in explaining cancer - the mitochondrial dysfunction theory offers a proximate explanation (the how), whereas the ancient gene reactivation theory offers an ultimate explanation (the why).
|
The what - cancer.
|